Прыжки по хромосоме
Введение
В последнее десятилетие мы стали свидетелями целой серии ошеломляющих успехов в области молекулярной биологии. Разработка надежных методов клонирования, секвенирования и анализа экспрессии эукариотических генов углубила наши представления о структуре и регуляции активности гена, сделала более понятными механизмы многих наследственных болезней человека. В это же время быстро развивались и достигли значительных успехов методы картирования человеческих генов.
До недавнего времени существовал некий «провал» в области размеров хромосомных сегментов от 100 до 5000 т. п.н.; для них не имелось адекватных методов исследования. Такое положение значительно усложняло интерпретацию данных по картированию, полученных методами генетики соматических клеток и с помощью генетического анализа. Сопряжение таких данных с информацией, полученной на молекулярном уровне, стали называть «обратной генетикой».
В последние годы было предложено несколько подходов, позволяющих вести исследования в этой новой области. В настоящей работе описан один из них – метод «прыжков по хромосоме». С его помощью удается клонировать последовательности ДНК, значительно удаленные на генетической карте от последовательностей, гомологичных используемому зонду. Излагаются методики создания библиотек «хромосомных прыжков» и клонов-связок. Обсуждены преимущества и недостатки описываемого метода.
1. Область применения метода
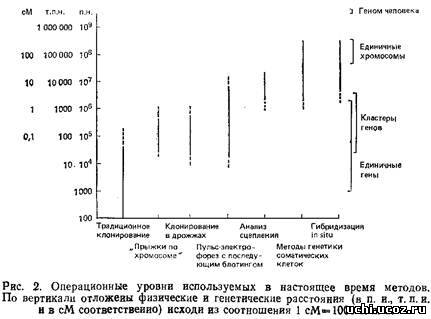
Рис. 2 дает представление о размерах хромосомных сегментов, в пределах которых «работают» различные современные методы генетических исследований. Ось ординат представляет собой логарифмическую шкалу физических расстояний, измеренных в парах нуклеотидов. На шкале приведены и значения генетических расстояний, измеряемые в сантиморганидах. 1 см приблизительно равна 108 п.н. Однако это соотношение нельзя считать универсальным, ибо зависимость между генетическим и физическим расстоянием на хромосоме имеет нелинейный характер, на нее могут оказывать влияние «горячие точки» рекомбинации. Наличие таких областей может привести к ситуации, когда сравнительно большому генетическому расстоянию соответствует небольшой отрезок на физической карте. В то же время в геноме существуют участки, рекомбинация в которых маловероятна, а это приводит к обратной ситуации. Как показано на рис. 2, классические методы молекулярной генетики хорошо работают на последовательностях длиной до 50 г.п.н., что соответствует максимальному размеру вставки в космидный вектор. Участки большей длины можно клонировать путем «прогулки по хромосоме», когда, используя уже клонированные последовательности, геномную библиотеку скринируют с целью получения перекрывающихся клонов. Таким способом удаётся анализировать последовательности длиной до нескольких сотен т. п.н. Однако, в некоторых случаях эта процедура может занять очень много времени. Так будет, если какие-то участки практически не перекрываются из-за наличия протяженной области повторяющихся последовательностей или последовательностей ДНК, которые не удается ввести в стандартные векторы. Если известно, что интересующий нас ген находится на расстояний нескольких сотен т. п.н. от используемого клона, то применение метода «прогулки по хромосоме» весьма проблематично.
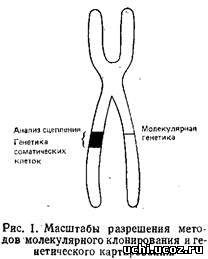
На противоположном конце спектра работают методы генетики соматических клеток, гибридизация in situ, анализ генетического сцепления; их разрешающая способность ограничена 1000–5000 т. п.н. И наконец, середине шкалы соответствуют три метода, позволяющие использовать данные картирования для поиска специфических молекулярных нарушений. Это пульс-электрофорез, «прыжки по хромосоме» и клонирование в клетках дрожжей.
Важность этих подходов в том, что они дают ключ к пониманию молекулярных основ целого ряда генетических нарушений, для которых неизвестна функция кодирующих их генов. Известны наследственные болезни человека, связанные с аномалиями отдельных генов, такие, например, как муковисцидоз, болезнь Гентингтона, нейрофиброматоз. Наследование их происходит строго в соответствии с менделевскими правилами, они имеют четкие фенотипические характеристики, однако нормальные функции генов, кодирующих эти заболевания, не определены. Анализ сцепления с использованием полиморфных ДНК-маркеров позволил картировать соответствующие гены в специфических хромосомах человека, что в свою очередь, значительно увеличило возможности пренатальной и пресимптоматической диагностики. Чтобы до конца разобраться в природе этих заболеваний и предложить адекватные методы их лечения, необходим молекулярно-биологический метод, работающий в области больших молекулярных размеров. Именно таким методом и являются «прыжки по хромосоме».
2. Стандартные библиотеки «прыжков»
2.1 Типы «прыжков»
Следует разграничить понятия стандартных геномных библиотек для «прыжков по хромосоме» и специфических библиотек. В первом случае библиотеки создаются таким образом, что начинать движение вдоль хромосомы отмеренными прыжками можно в принципе с любой точки генома. Специфические библиотеки состоят из клонов, позволяющих осуществлять прыжки от одного редко встречающегося сайта рестрикции, например Notl, к последующему такому же сайту. Типы прыжков схематически изображены на рис. 3. Очевидно, что способы создания этих библиотек несколько различаются. С технической точки зрения труднее получать библиотеки первого типа, так как они должны содержать репрезентативную выборку последовательностей генома. Обычно для такой библиотеки требуется 3*108 клонов. Тогда можно быть твердо уверенным, что с ней можно работать, начиная от любой стартовой точки на хромосоме.
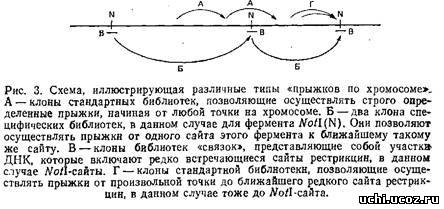
Для создания же полных специфических библиотек требуется всего лишь 10000-20000 клонов, так как количество независимых клонов эквивалентно числу рестрикционных фрагментов, полученных при использовании данного фермента рестрикции. Так, например, для рестриктазы Notl, которая отщепляет приблизительно по 1000 т. п.н. в геноме человека, таких фрагментов должно быть всего лишь около 3000. Поэтому библиотеку из 10000 клонов для прыжков по Not сайтам можно считать практически полной. Очевидным недостатком таких библиотек является то, что их нельзя использовать, когда стартовая точка прыжка не примыкает к редко встречающемуся сайту рестрикции. К сожалению, это довольно частое явление, препятствующее быстрому распространению данного метода. Если же все-таки удается идентифицировать клон, примыкающий к редко встречающемуся сайту рестрикции, использование специфических библиотек для «прыжков по хромосоме» может оказать неоценимую помощь.
Очень эффективным могло бы оказаться создание геномной библиотеки третьего типа, имеющей в своем составе клоны, соединяющие редко встречающиеся сайты рестрикции с прилегающими случайными последовательностями ДНК генома. Имея такую библиотеку, мы могли бы начинать движение по хромосоме с любой стартовой точки, а при встрече с редким сайтом рестрикции пускать в ход специфическую геномную библиотеку. К сожалению, пока эта задача не решена и подходы к ее решению здесь обсуждаться не будут.
2.2 Принцип создания стандартных библиотек
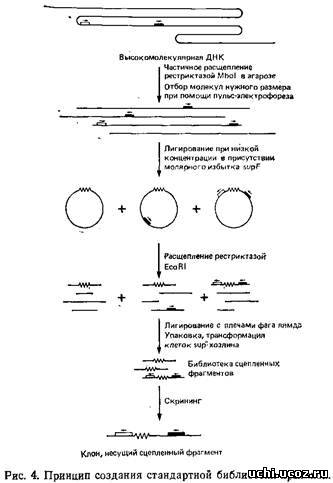
Основная стратегия метода «прыжков по хромосоме» состоит в получении кольцевых формул очень крупных фрагментов ДНК путем лигирования их при большом разбавлении. Образование колец позволяет физически сблизить участки ДНК, расположенные в геноме на значительном расстоянии друг от друга. Селективное клонирование таких соединенных фрагментов в стандартные векторы позволяет затем получить геномную библиотеку клонов – «прыжков». Эта стратегия применительно к стандартным библиотекам схематически изображена на рис. 4.
Прежде чем приступить к созданию библиотеки, следует ответить на несколько важных вопросов.
1. Какой размер прыжка желателен? Известно, что размер прыжка определяется размером частично гидролизованных молекул ДНК. Как будет показано ниже, сложность построения библиотеки возрастает в степени 3/2 с увеличением размера прыжка.
2. Какой использовать фермент? В идеальном случае хотелось бы иметь совершенно случайный набор фрагментов ДНК.
3. Какой источник ДНК использовать? Для оптимальной реализации библиотеки при наличии самых разнообразных стартовых зондов было бы предпочтительнее использовать источник ДНК, представляющей весь геном организма, например периферические лимфоциты донорской крови в случае геномных библиотек человека. Если же предполагается исследование специфической хромосомы, лучше работать с гибридными соматическими клетками, содержащими именно эту хромосому на известном фоне хромосом других видов. Преимущество такой стратегии в том, что она позволяет сразу определить принципиальную ценность клона, полученного при помощи имеющейся библиотеки, путем простой проверки клонированного фрагмента на его принадлежность интересующей хромосоме.
2.3 Получение ДНК-фрагментов желаемого размера
ДНК, подвергаемая частичному гидролизу, должна быть достаточно высокомолекулярной. Методика приготовления образцов ДНК та же, что и для пульс-электрофореза, с той лишь разницей, что количество ДНК рассчитывается для препаративных целей.
1. Исходя из того, что одна клетка млекопитающего содержит 6,7 пкг ДНК, необходимо вырастить их столько, чтобы получить достаточное количество ДНК. Обычно для создания библиотеки требуется 200 мкг ДНК, что соответствует приблизительно 3х107 клеткам. ДНК должна быть очень высокого качества, поэтому очень важно растить клетки в оптимальных условиях. Можно приготовить клетки и из периферической крови центрифугированием в смеси Ficoll-Hypaque.
2. После сбора клеток аккуратно просчитайте их количество в гемоцитометре и суспендируйте в таком объеме фосфатно-солевого буфера, чтобы концентрация клеток составляла примерно 2хЮ7/мл. Затем суспензию быстро смешайте с равным объемом расплавленной 2%-ной низкоплавкой агарозы в 125 мМ ЭДТА при 40°С. Залейте в форму. Кроме стандартных ячеек форма должна иметь ячейки для блоков размером 2x8x135 мм, которые служат для получения ДНК в препаративных количествах. Полезно приготовить 10–20 стандартных блоков с ДНК для тест-гидролизатов и два больших блока непосредственно для опытов.
3. Обработайте ДНК протеиназой К в присутствии больших количеств ЭДТА и N-лаурилсаркозина, как описано ранее.
4. Очищенную ДНК проверьте на нативность и на присутствие нуклеаз. Для этого инкубируйте половину содержимого блока при 37°С 3 ч с 10 мм MgCl2 и нанесите эту смесь, а также необработанную половину на OFAGE-гель или на гель для электрофореза в инвертированном поле. Определите размер ДНК. Необработанная ДНК должна практически полностью остаться в лунке, а в обработанном магнием препарате не должно быть низкомолекулярных примесей. Только в этом случае можно использовать препараты для «прыжков по хромосоме». Если же в препаратах, обработанных магнием, налицо признаки деградации, значит они загрязнены нуклеазами и надо повторить обработку протеиназой К. Как правило, этого бывает достаточно, чтобы избавиться от загрязнений.
5. Из некоторых источников ДНК постоянно выделяется с небольшим количеством низкомолекулярных примесей размером 50–100 т. п.н., видимо из мертвых клеток. Наиболее характерно это для лимфобластов. Такие деградировавшие молекулы ДНК могут сильно искажать результаты «прыжков по хромосоме», поэтому необходимо удалить их из агарозных блоков перед обработкой рестриктазами. Для этого блоки помещают в лунки OFAGE-геля и проводят пульс-электрофорез в течение 2–3 ч с интервалами между импульсами 20 с. Молекулы ДНК размером менее 100 т. п.н. выходят из блоков в гель, а высокомолекулярная ДНК остается практически без изменения. Блоки затем можно изъять из геля, получив, таким образом, высококачественный материал для дальнейшего исследования.
6. Проведите контрольную рестрикцию половинок агарозных блоков различными концентрациями Mbol. Согласно данной методике, в каждой половине блока содержится примерно 3,3 мкг ДНК, следовательно, концентрация фермента составит от 0,01 ед./мкг до 0,045 ед./мкг. Остановите реакцию добавлением 10 мкл ЭДТА. Обработанные рестриктазой образцы поместите в гель и проведите пульс-электрофорез. Это поможет определить концентрацию фермента, оптимальную для получения фрагментов нужного размера. На рисунке наглядно продемонстрирована необходимость пре-электрофореза для удаления низкомолекулярных примесей ДНК перед обработкой рестриктазами.
7. При использовании больших агарозных блоков пропорционально увеличьте количество фермента, но так, чтобы концентрации фермента в ед./мкл и ДНК в мкг/мл остались прежними.
8. Обработанные рестриктазой препаративные блоки поместите в гель и, используя соответствующие маркеры, проведите пульс-электрофорез с интервалами между импульсами, позволяющими хорошо отделить фрагменты нужной величины.
Если используется OFAGE-гель, маркеры следует наносить в середине и по обоим краям геля. Мы обнаружили, что для получения прямых полос молекул ДНК желаемых размеров лучше использовать CHEF-гели и гель-электрофорез в инвертированном поле.
9. Важно, чтобы ДНК, которую предстоит клонировать, не подвергалась ультрафиолетовому облучению. Поэтому отрежьте края геля, содержащие маркеры, а также небольшое количество рестрицированной геномной ДНК, окрасьте их бромидом этидия и визуализируйте в ультрафиолете. По полученным данным определите участок геля с интересующей вас ДНК.
10. Вырежьте нужный участок геля. На этой и последующих стадиях важно использовать безнуклеазные реактивы и инструменты. Выделять ДНК из геля для получения кольцевой молекулы можно двумя способами. Нам представлялось удобным делать это методом эле